Tous droits réservés © NeurOreille (loi sur la propriété intellectuelle 85-660 du 3 juillet 1985). Ce produit ne peut être copié ou utilisé dans un but lucratif.
 Français
Français
 English
English
 Español
Español
 Português
Português
Outer hair cells (OHCs) are disposed in three rows on the external side of the tunnel of Corti. OHCs are quite atypical sensory cells. They start transduction classically (as IHCs), being excited by K+ entrance into the tips of stereocilia. However, this excitation insteed of initiating an auditory message, triggers a reverse transduction process, feeding energy back into the Corti's organ. This electro-mechano transduction (also called active mechanism), due to OHC electro-motile properties, enhances the cochlear sensitivity and frequency selectivity. The human cochlea bears ten to twelve thousands OHCs.
Schematic of an outer hair cell (OHC)
")
OHCs have a regular cylindric and elongated shape. At the synaptic pole, two types of connections are seen: an efferent synapse between a medial efferent terminal (red) and the OHC, and an afferent synapse between the OHC and spiral afferent (green).
")
")
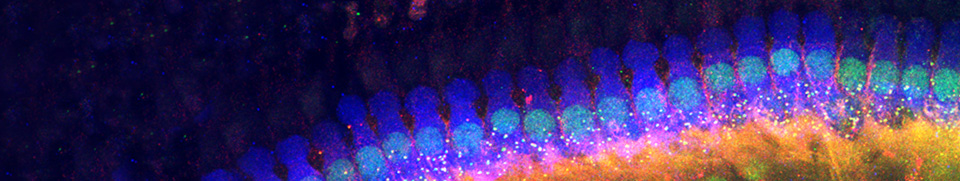
Arrangement of OHC stereocilia

M.Lenoir |

R Pujol |
|
| Staircase arrangement of the 3 rows of OHC stereocilia (SEM, rat cochlea, basal turn) Note the typical "W" arrangement. The inside cuticular plate is devoid of microvilli, which cover the surface of surrounding Deiters cells. Scale bar: 1 µm |
Enlargement of the cuticular plate of an OHC from the above picture. The plane of section allows a precise view of the "W" pattern and a count of the 3 rows of stereocilia. Also, the tight coupling with adjacent Deiters' cells is clearly visible ; this seals the junction between endolymphatic and perilymphatic (spaces of Nuel) compartments. Scale bar: 2 µm |
OHC length variation according to frequency (the "xylophone")
Remarkably, the length of OHCs varies according to the place of frequency coding. This is true within one species and accross species. Up to know, no explanation of this interesting feature has been proposed. Is that related to some physical properties such as resonance ?

Schematic drawing representing OHCs from different mammalian species and different cochlear turns. While OHC diameter keeps a constant value (7 µm), their length regularly varies according to frequency. In the human cochlea, a 25 µm basal OHC (C) is found at a place which codes for 20 kHz; conversely a 70 µm OHC (G) is found apically at the site coding for a very low frequency (< 100 Hz). A = shortest OHC in basal turn of a bat cochlea (at a place coding for 160 kHz), B = basal OHC from a cat cochlea (at a place coding for 40 kHz), D = OHC from second turn of a guinea pig cochlea (at a place coding for 5 kHz), E = OHC from start of third turn of a guinea pig cochlea (at a place coding for 2.5 kHz), F = OHC from end of third turn of a guinea pig cochlea (at a place coding for 150 Hz), H = apical OHC from a mole rat cochlea (at a place coding for 15 Hz).
Facebook Twitter Google+