Cochlea: function
Authors: Guy Rebillard, Rémy Pujol
Contributors: Sam Irving
The cochlea is capable of exceptional sound analysis, in terms of both frequency and intensity.
The human cochlea allows the perception of sounds between 20 Hz and 20 000 Hz (nearly 10 octaves), with a resolution of 1/230 octave (from 3 Hz at 1000 Hz).
At 1000 Hz, the cochlea encodes acoustic pressures between 0 dB SPL (2 x 10-5 Pa) and 120 dB SPL (20 Pa).
The transfer of airborne sound vibration to the structures and fluid of the cochlea
When sound pressure is transmitted to the fluids of the inner ear by the stapes, the pressure wave deforms the basilar membrane in an area that is specific to the frequency of the vibration. In this way, higher frequencies cause movement in the base of the cochlea, and deeper frequencies work at the apex. This characteristic is known as cochlear tonotopy.
High: 10kHz
Low: 1kHz
Animations by S. Blatrix and G. Rebillard
Frequency distribution along the basilar membrane of a human cochlea: passive tonotopy

A few characteristic frequencies (in kHz) are shown in blue. Note the gradual thickening of the basilar membrane from the base (20kHz) to the apex (20Hz).
A brief history of frequency coding
Throughout history, numerous theories have been suggested to explain frequency coding.
Resonance
In the middle of the 19th century, Ludwig von Helmholtz suggested that resonators tuned to different audible frequencies were spread along the basilar membrane. He suggested a role for the pillars of Corti and a system of fibres stretched over the basilar membrane like piano strings.
Note that after having been superceded for a few decades by the “travelling wave” theory, this first suggestion returns in another form. A number of acousticians today think that the most realistic model of basilar membrane function is the resonator system, or, even better, a system of frequency-tuned oscillators that can be regulated by the central nervous system (known as efferent feedback).
Travelling wave

George Von Bekesy (winner of the 1961 Nobel prize for physiology and medicine) distanced himself from resonance theory from the 1950s. Using cochleae taken from human cadavers and hydraulic models, he demonstrated that when the cochlea was stimulated by a sound wave, the basilar membrane motion was a wave that travelled from the base of the cochlea.
The amplitude of this wave increased with distance travelled, arrived at a maximal point and then experienced a rapid drop off. The point of maximal amplitude of the travelling wave depended upon the sound frequency: higher sounds had maximal points at the base of the cochlea, and lower sounds towards the apex (see the animation at the top of the page).
Active mechanism
The theory of the travelling wave, and the mechanical filtering of frequencies, was quickly shown to be too simplistic to explain the excellent frequency discrimination of the cochlea.
It was only at the end of the 1990s that Johnstone and Boyle showed, in living animals, that the vibration of the basilar membrane was greater and more focalised than Bekesy had suggested.
This discovery helped to explain the cochlea’s frequency selectivity. We now know that the differences between Bekesy and Johnston’s observations were due to active biological mechanisms that act upon the vibration of the basilar membrane in living subjects (see below).
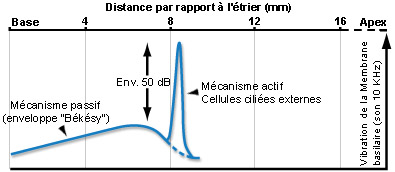
For a pure tone sound, active mechanics amplify basilar membrane vibrations by around +50 dB at a very narrow section of the organ of Corti, which serves to increase the sensitivity of the cochlea at this site. Two similar frequencies can therefore activate two distinct cochlear regions, allowing them to be differentiated (a characteristic known as frequency selectivity). This frequency tuning is closely linked to the electromotility of the outer hair cells (OHCs), and is defined by the fibres of the auditory nerve and the inner hair cells (IHCs) that generate the neural signal (see OHCs physiology).